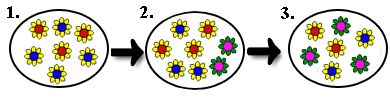
Es gibt Eigenschaften, die sich auf den ersten Blick schwer mit der Evolutionstheorie der natürlichen Selektion erklären lassen. Insbesondere bei Verhaltensweisen scheint es unlogisch, weshalb ein Organismus sich für andere opfern soll, indem beispielsweise ein Murmeltier durch Pfeifen andere auf eine drohende Gefahr hinweist und sich so aber selber grossen Risiken aussetzt, oder weshalb bestimmte Lebewesen ganz auf Fortpflanzung verzichten (beispielsweise die Arbeiterinnen bei den Bienen). Dieses selbstlose Verhalten, oder Altruismus, scheint im Widerspruch zu stehen mit der Idee, möglichst lange zu überleben, um möglichst viele Nachkommen zu zeugen. „In der Evolutionsbiologie wird mit dem Begriff Altruismus oder Selbstlosigkeit kein absichtliches Handeln verbunden. Vielmehr versteht man unter Altruismus Verhaltensweisen eines Individuums, von denen überwiegend andere Individuen im Sinne eines im Vergleich zum Altruisten relativ höheren Fortpflanzungserfolgs profitieren. Der Begriff definiert sich hier also über seine Konsequenzen hinsichtlich des relativen individuellen Fortpflanzungserfolges der Beteiligten und nicht über eine „Absicht“ (die jedoch zusätzlich vorliegen kann, vor allem beim Menschen). Altruismus scheint auf den ersten Blick dem Darwin'schen Prinzip zu widersprechen, wenn z. B. ein Tier unter Energieaufwand einem Artgenossen hilft, ohne davon irgendwelche Vorteile zu gewinnen. Dies würde einem Individuum aus energetischer Sicht einen Nachteil verschaffen. Allerdings kann dieser Nachteil dadurch wieder ausgeglichen werden, wenn der Population oder Art dadurch Vorteile entstehen.“
Die Evolution kann mit Modellen beschrieben werden; sogar Verhaltensweisen gehorchen mathematischen Gleichungen. 1968 beschrieb William D. Hamilton einen Zusammenhang zwischen Verwandtschaftsgrad und „Hilfsbereitschaft“:
„Die Gesamtfitness eines Lebewesens kann demnach als die Anzahl der eigenen Gene, die an die nachfolgende Generation weitergegeben wird, gemessen werden. Nach John Maynard Smith setzt sie sich zusammen aus
i) der direkten Fitness, den eigenen Genen in den eigenen Nachkommen, und
ii) der indirekten Fitness, den eigenen Genen, die durch Verwandte zusätzlich an fremde Nachkommen weitergegeben wurden.
Da Verwandte zum Teil dieselben Gene besitzen wie das Individuum, fördert dieses durch Helferverhalten die Weitergabe des eigenen Erbguts (Verwandtenselektion). Dieser Altruismus ist nur dann erfolgreich und breitet sich aus, wenn der Nutzen für denjenigen, der das altruistische Verhalten zeigt, grösser ist als die Kosten, die er dafür investieren muss (Hamiltons Regel).
Mathematisch ausgedrückt muss das Verhältnis von Nutzen (B) zu Kosten (C) grösser sein als eins dividiert durch den Verwandtschaftsgrad:
B/C > 1/r beziehungsweise r x B > C, mit B: Nutzen (benefit); C: Kosten (cost); r: Verwandtschaftskoeffizient (relatedness).
Beispiel: Ein Tier, das durch seine Hilfe auf zwei eigene Nachkommen verzichtet (C = 2), dafür aber einem Geschwister (Verwandtschaftsgrad zwischen Geschwistern bei diploiden Organismen (r = 0,5) hilft, fünf zusätzliche Nachkommen (B = 5) zu produzieren, hat eine höhere Gesamtfitness als ein Tier, das „egoistisch“ nicht hilft.“ (Quelle: Wikipedia).
Der Verwandtschaftskoeffizient ergibt sich entsprechend der Vererbungslehre nach dem folgenden Schema.
Die physikalische Evolution beschreibt den Anfang von allem, nämlich der Entwicklung des Weltalls. Diese beginnt mit dem Urknall. Dabei wurde spontan so viel Materie frei, dass sich diese heute noch durchs Universum bewegt, mit der Antriebsenergie von vor Milliarden Jahren. Das Universum wird, so vermuten Forscher, auch noch weiter wachsen, sich also ausdehnen. Darauf folgte die Entstehung von Subteilchen (Elementarteilchen) wie Quarks und Antiquarks. Diese bestehen aus Energie. Daraus bildeten sich die ersten Atome, also Teilchen (Materie). Daraus wiederum entstanden die ersten Moleküle und andere Ketten.
Quelle: www.helpster.de
2. (Bio-)Chemische Evolution und die Entstehung des Lebens auf der Erde
Die chemische Evolution im weitesten Sinne beginnt bereits mit der Synthese von schwereren chemischen Elementen durch Kernfusionsprozesse (Nucleogenese) im Innern der Sterne (nur Wasserstoff, Helium und Spuren von Lithium, Beryllium und Bor wurden in den ersten Minuten unseres Universums mit dem Urknall erzeugt). Vom Wasserstoff abgesehen, sind also alle für das irdische Leben essentiellen Elemente in Sternen "erbrütet" worden. Die durch Sternwinde und -explosionen ins All abgegebenen Elemente stehen als Rohstoffe für die Entstehung neuer Sterne sowie Planeten zur Verfügung, können sich aber auch im interstellaren Raum selbst zu Molekülen verbinden, z. B. zu Wasser, Kohlendioxid, Methan usw., aber auch zu einfachen Aminosäuren wie Glycin und sogar zu langkettigen Kohlenwasserstoffen, insbesondere den polycyclischen aromatischen Kohlenwasserstoffen (PAHs). Rund 150 verschiedene Molekülsorten wurden im Weltraum bereits spektroskopisch nachgewiesen.
Die chemische Evolution im engeren Sinne ist die Entstehung von Biomolekülen auf der Urerde. Diese Moleküle waren und sind die chemische Grundlage des (irdischen) Lebens. Dazu gehören insbesondere die Ribonucleinsäuren und Proteine (bzw. Polypeptide) und ihre Bausteine. Die Uratmosphäre war nach einer gängigen Auffassung reduzierend. Sie enthielt keinen Sauerstoff, aber neben Wasser, Stickstoff und Kohlendioxid kleinere Anteile von Kohlenmonoxid, Methan, Ammoniak, Schwefeldioxid und möglicherweise Wasserstoff. In den zwanziger Jahren entwickelten A.I. Oparin und J.B.S. Haldane unabhängig voneinander die Hypothese, daß durch die ultraviolette Strahlung der Sonne, die heute größtenteils von der Ozonschicht in den oberen Bereichen der Atmosphäre absorbiert wird, oder durch elektrische Entladungen (Blitze) und andere Energiequellen einfache organische Verbindungen wie Aminosäuren, Nucleinsäurebasen und Zucker aus den Molekülen der Uratmosphäre entstanden seien. Daß dies tatsächlich möglich ist, haben 1953 S.L. Miller (Miller-Experiment) und H.C. Urey experimentell erwiesen.
Diese abiotischen Synthesen produzierten neben unlöslichem Teer beträchtliche Mengen wasserlöslicher organischer Verbindungen, darunter zahlreiche Aminosäuren (die Bausteine der Proteine) und andere biochemisch bedeutsame Komponenten wie Formaldehyd und Cyanwasserstoff (Blausäure). Weitere Experimente mit veränderten Reaktionsbedingungen, Gasgemischen oder Energiequellen führten zur Synthese weiterer Aminosäuren sowie von Zuckern und Nucleinsäurebasen. Die Bildung von Zuckern könnte aus der Kondensation von Formaldehyd mit Hilfe zweiwertiger Kationen, Erden oder Tonen als Katalysatoren erfolgt sein. Nucleinsäurebasen (insbesondere Adenin) entstehen aus der durch Ammoniak katalysierten Kondensation von Cyanwasserstoff. Uracil kann sich durch die Einwirkung energiereicher Protonen aus Kohlenmonoxid, Stickstoff und Wasser bilden, Cytosin sowohl aus Cyanoethan und Cyanat als auch aus Cyanoacetaldehyd und Harnstoff oder aus Methan, Stickstoff, Ammoniumionen und Wasser.
Vermutlich ist es kein Zufall, daß die so entstandenen Verbindungen die Grundbausteine der Biomoleküle darstellen. Sie kamen in präbiotischer Zeit wahrscheinlich relativ häufig im Urozean vor. Nach einer Hypothese von der Entstehung des Lebens sind in Pfützen im Uferbereich infolge der katalytischen Wirkung bestimmter Mineralien Polypeptide und Polynucleotide (Nucleinsäuren) kondensiert, aus denen dann durch rückgekoppelte Prozesse einer molekulardarwinistischen Selbstorganisation die ersten Lebensformen entstanden sind.
Ingredienzien für die Ursuppe:
Obwohl reduzierende Atmosphären noch heute im Sonnensystem existieren, etwa bei Jupiter, Saturn und dessen Mond Titan, kamen in jüngerer Zeit Zweifel auf, ob auch die Urerde wirklich eine solche Atmosphäre besaß. Gesteinsuntersuchungen und verschiedene theoretische Überlegungen zur Erdentstehung lassen es möglich erscheinen, daß die Uratmosphäre neutral war und vorwiegend aus Kohlendioxid, Stickstoff und Wasser bestand. Dann wären die Bedingungen für die Synthese der präbiotischen Verbindungen allerdings ungünstiger gewesen. Nach C.F. Chyba, J.M. Greenberg, J. Oro, C. Sagan und anderen könnten die mitunter als Ursuppe bezeichneten Urmeere allerdings durch Ingredienzien aus dem All angereichert worden sein. Dafür spricht, daß sowohl in den interplanetaren Staubteilchen, die man in letzter Zeit auffangen konnte, als auch in Kometen große Mengen an Kohlenstoff-Verbindungen vorkommen (bis zu 20% der Masse). Noch heute rieseln jährlich rund 40 000 Tonnen Staub aus dem All auf die Erde. In Meteoriten wurden ebenfalls zahlreiche organische Moleküle gefunden – insbesondere Aminosäuren, die den Flug durch die Atmosphäre zumindest teilweise überstanden. Laborexperimenten zufolge können sich Aminosäuren und andere organische Moleküle auch bilden, während Meteoriten durch methanhaltige Atmosphären stürzen.
Der Beginn der Stammesgeschichte des Menschen, lässt sich etwa zu dem Zeitpunkt verorten, an dem die letzte Aufspaltung einer zuvor gemeinsamen Population von Vorfahren des Menschen und der Schimpansen stattfand. Schimpansen sind dem Menschen auf phylogenetischer Basis am nahesten. Das menschliche Erbgut unterscheidet sich nur zu 1,37 % von dem des Schimpansen, zu 1,75 % vom Gorilla und zu 3,4 % vom Orang-Utan (Quelle: www.biologie-schule.de).
Erste Hominiden (Menschenartige)
Gegen Ende der
erdgeschichtlichen Epoche des Tertiär
vor rund 3-4 Millionen Jahren traten die
ersten Urmenschen(Australopithecus) auf. Skelettfunde in
Äthiopien und Australien belegen, dass sie mit 500 cm³
ein nur wenig grösseres Gehirn als heutige
Menschenaffen hatten, aber schon aus Steinen einfachste Werkzeuge
herstellten und aufrecht gingen. Die neuere Forschung geht davon aus,
dass die Menschen nicht von den heutigen Menschenaffen abstammen,
sondern gemeinsame Vorfahren in Ostafrika hatten, die ausgestorben
sind.
Homo habilis und homo erectus
Homo habilis
Etwas weiter entwickelt als der Australopithecus war der Homo habilis vor rund 2 Millionen Jahren mit einem Gehirnvolumen von 600 - 700 cm³, der sich vor rund 1.5 Millionen Jahren zum Homo erectus mit einem Gehirnvolumen von rund 800 cm³ weiterentwickelte. Der homo erectus lebte in Afrika, Asien und Europa und war bereits in der Lage, auf dem Feuer zu kochen.
Altsteinzeit: Frühmenschen, Altmenschen und der Homo sapiens
In den letzten Teil der erdgeschichtlichen Periode des Quartär, nämlich in die Eiszeiten, fällt das Auftreten der
Frühmenschen: (Homo erectus) vor rund 500'000 Jahren
Javamensch (Pithecanthropus, Homo erectus erectus) Gehirnvolumen 775-950 cm³
Pekingmensch (Sinanthropus, Homo erectus pekinensis) Gehirnvolumen 900-1100 cm³, benutzte bereits das Feuer
Heidelbergmensch (Homo erectus heidelbergensis)
Altmenschen (Homo sapiens)
Die Entwicklungslinie des Menschen spaltete sich in der zweiten grossen Zwischeneiszeit vor rund 300'000 - 200'000 Jahren auf in
die Neandertaler (Homo sapiens neanderthalensis), Gehirnvolumen 1440 cm³, die recht gut an das kältere Klima der letzten Eiszeit angepasst waren, bereits vielfältige Werkzeuge und Waffen besassen, aber um 26000 v. Chr. in Südeuropa wieder ausgestorben sind und
die Jetztmenschen (Homo sapiens sapiens)
Jetztmenschen (Homo sapiens sapiens)
Wie die Entwicklung in den Eiszeiten verlief, ist unklar. Die ersten aus Funden bekannten Vertreter der modernen Menschen sind die Cromagnon-Menschen (vor rund 40'000 Jahren) mit einem Gehirnvolumen von 1400 cm³.
Die Cromagnon Menschen
Sie bemalten Höhlenwände mit Naturfarben (wahrscheinlich zur Beschwörung der Beutetiere; von rund 120 Fundorten sind Altamira in Nordspanien und Lascaux in Frankreich die Bedeutendsten. Daneben in der Schweiz Schweizersbild SH (entdeckt 1893) und Kesslerloch SH.
Die Cromagnon Menschen stellten Schmuck und religiöse Kunstwerke her.
Alle heutigen Menschen gehören zur Art Homo sapiens sapiens und stammen nach neusten Erkenntnissen der Genforschung von einer relativ kleinen Gruppe ab, die in Afrika lebte. Diese Periode wird in der Entwicklung der Menschen als Altsteinzeit (Paläolithikum) bezeichnet.
Als Arten bezeichnet man Gruppen von Individuen, die durch Abstammungsbande zwischen Elter(n) und Nachkommen (Nachkommenschaft) gekennzeichnet sind
und in Gestalt, Physiologie und Verhalten soweit übereinstimmen, daß
sie sich von anderen Individuengruppen abgrenzen lassen. Bei Organismen
mit zweigeschlechtlicher Fortpflanzung kommt als entscheidendes Kriterium die Fähigkeit hinzu, gemeinsam fertile Nachkommen
zu erzeugen. Die Angehörigen einer Art bilden demnach eine Fortpflanzungsgemeinschaft, zwischen ihnen besteht Genfluß, sie haben Anteil an einem Genpool und sind somit die Einheit, in der evolutionärer Wandel stattfindet (Evolution). – In aller Regel lassen sich die Individuen verschiedener Biospezies anhand äußerer Merkmale
unterscheiden, doch liefern nicht diese, sondern allein die zu
beobachtenden Fortpflanzungsbeziehungen das entscheidende Kriterium
artlicher Zuordnung. Dies zeigt sich vor allem in kritischen Fällen, z.
B. bei stark ausgeprägtem Sexualdimorphismus
(wo man die beiden Geschlechter primär unterschiedlichen Arten zuordnen
würde) oder bei den morphologisch sehr ähnlichen, aber genetisch
isolierten Zwillingsarten (die man fälschlich als eine Art auffassen kann) (Quelle: www.spektrum.de).
Reproduktive Isolation
Die reproduktive Isolation bildet die essentielle Grundlage der Artentstehung. Bei einer reproduktiven Isolation besteht durch eine oder mehrere Fortpflanzungsbarrieren kein Genfluss mehr.
Fortpflanzungsbarrieren können innerartlich, aber auch außerartlich wirken und führen dauerhaft zum Entstehen einer neuen
Art (Ausnahme: Isolation durch Sterilität).
Man unterscheidet mehrere Formen von Isolation:
Zeitliche Isolation
Arten können sich nicht miteinander fortpflanzen, weil sie sich während unterschiedlicher Jahreszeiten/Tageszeiten fortpflanzen
(z.B. bei Froscharten, deren Paarungszeit in unterschiedlichen Monaten liegt)
Habitat Isolation
Zwei eng verwandte Arten von Strumpfbandnattern paaren sich nicht, da eine Art an Land lebt während die Andere hauptsächlich im Wasser.
Durch zufällige Mutationen können Individuen nicht mehr mit der
Ursprungspopulation fortpflanzungsfähig sein.
Physiologische Isolation
Aufgrund der unterschiedlichen Form der Kopulationsorgane können sich bestimmte Arten nicht miteinander fortpflanzen
(z.B. bei nah verwandten Insektenarten). Bei den hier gezeigten verschiedenen Schneckenarten drehen die Schneckenhäuser in entgegengesetzten Richtungen. Hierdurch können die Genitalöffnungen )siehe Pfeile) nicht in die richtige Position gebracht werden und somit keine Paarung durchgeführt werden.
Physiologische Isolation bei Schnecken
(Quelle: http://wps.pearsoncustom.com)
Ökologische Isolation
Durch das Ausnutzen von unterschiedlichen ökologischen
Nischen im selben Gebiet kommt es zu einer reproduktiven Isolation (z.B. Darwinfinken, die unterschiedliche ökologische Nischen
besetzten; manche Individuen ernährten sich von Insekten, andere von Körnern).
Ökologische Isolation bei Darwinfinken
Geografische Isolation
Wegen geografischen Barrieren können Teilpopulationen einer Art sich nicht untereinander fortpflanzen
(z.B. durch Kontinentaldrift oder eine Erhöhung des Meeresspiegels).
Verhaltensisolation
Unterschiedliches Verhalten während der Paarungszeit isoliert Arten voneinander (z.B. haben nah
verwandte Vogelarten, die sich zur selben Zeit paaren, einen unterschiedlichen Balzruf).
Sterilität
Bei der Kreuzung zweier nicht verwandten Arten können Bastarde mit einem ungeraden Chromosomensatz entstehen, die nicht
Fortpflanzungsfähig sind (z.B. bei einer Kreuzung aus Esel und Pferd sind die Nachkommen allesamt unfruchtbar, weil sie einen
ungeraden Chromosomensatz besitzen, der eine Bildung von Gameten unmöglich macht).
Die Kreuzung von Pferd und Esel bringt Nachkommen mit einem ungeraden Chromosomensatz hervor, die nicht fortpflanzungsfähig sind.
(Quelle: http://wps.pearsoncustom.com)
Polyploidie
Unter Polyploidie versteht man das Vorhandensein von mehr als zwei Chromosomensätzen. Fortpflanzung ist nur unter Individuen mit
identischer Anzahl von Chromosomensätzen möglich. (z.B. bilden Pflanzen häufig triploide oder tetraploide Chromosomensätze und
sind somit von ihrer Ursprungsart mit haploidem Chromosomensatz genetisch isoliert).
(Quelle: www.biologie-heute.de,
http://wps.pearsoncustom.com)
Fortpflanzungsbarrieren als Resultat der Isolierung
Bei einer Isolation können sich Fortpflanzungsbarrieren entwickeln. Die zeigt das folgende Experiment von Diane Dodd: Eine Population von Fruchtfliegen wurde aufgeteilt und fortan für mehrere Generationen entweder in Anwesenheit von Maltose oder Stärke als Kohlenstoffquelle kultiviert. Anschliessend wurde ein Paarungsexperiment durchgeführt, und die Paarungsversuche statistisch ausgewertet. Das Experiment zeigt, dass sich die Fliegen, die Stärke als Kohlenstoffquelle gewohnt sind, auch präferentiell mit solchen Fliegen paaren. Ebenso verhalten sich die Fliegen, die Maltose als Kohlenstoffquelle gewohnt sind. Zwar treten auch Paarungen zwischen den Gruppen auf (Stärke-Maltose) , jedoch sind diese deutlich seltener (< 50%) .
Diane Dodd: Als Folge von isolation entstehen Fortpflanzungsbarrieren (Quelle: http://wps.pearsoncustom.com)
Isolation von Populationen kann zu Fortpflanzungsbarrieren führen (Quelle: UC Berkeley)
Sympatrische Artenbildung
Die Entstehung einer neuen Art im Gebiet der Ursprungsart nennt man sympatrische Artbildung. Bei dieser Form der Artbildung
muss es, beispielsweise durch Polyploidisierung bei Pflanzenarten, zu
einer unmittelbaren reproduktiven Isolationen kommen, damit
die Pflanze keinen Allelaustausch mehr mit der übrigen Population hat.
1) Sympatrische Artenbildung durch Polyploidisierung
Viele Pflanzenarten sind das Ergebnis eines Fehlers bei der Zellteilung, durch welchen ein zusätzlicher Satz an Chromosomen entstanden ist. Arten, die sich auf diese Weise gebildet haben sind polyploid, d.h. ihre Zellen haben mehr als zwei Chromosomensätze. In der folgenden Abbildung ist gezeigt, wie sich aus einer diploiden Pflanze (2n) eine tetraploide Pflanze entwickeln kann:
Sympatrische Artenbildung durch Polyploidisierung (Quelle: http://wps.pearsoncustom.com)
1)
Ein Fehler bei der Zellteilung führt nach der Replikation des
diploiden Chromosomensatzes (2n) zu einer Verdopplung der Chromosomen.
2)
Wenn sich aus diesen tetraploiden Zellen (4n) eigenständige
Pflanzen entwickeln, bilden diese Pflanzen Blüten mit diploiden Gameten
(Keimzellen) aus.
3)
Wenn diese Pflanzen sich selbst befruchten (was häufig bei Pflanzen
vorkommt), entwickeln sich aus den tetraploiden Zygoten erneut
tetraploide Pflanzen, die fruchtbar sind und durch Selbstbefruchtung
oder Befruchtung durch andere tetraploide Pflanzen neue, tetraploide
Nachkommen bilden. Die so entstandenen tetraploiden Pflanzen können jedoch keine fruchtbaren Nachkommen erzeugen, wenn sie durch die Elternpflanzen befruchtet werden (und auch umgekehrt nicht !). Die Verschmelzung der diploiden Keimzellen der tetraploiden Pflanze mit den haploiden Keimzellen der diploiden Elternpflanze würde eine triploide Nachkommenschaft (3n) erzeugen. Triploide Individuen sind jedoch unfruchtbar. Sie können keine normalen Keimzellen bilden, da durch die ungerade Chromosomenzahl während der Meiose keine homologen Chromosomenpaare gebildet werden können, die sich anschliessend durch Zellteilung gleich aufteilen würden. Somit wird durch die Bildung eines tetraploiden (4n) Pflanzennachkommens innerhalb nur einer Generation eine neue Art geschaffen, die reproduktiv isoliert ist.
2) Sympatrische Artenbildung durch Hybridisierung
Die meisten polyploiden Pflanzen entwicklen sich jedoch durch Hybridisierung zweier unterschiedlicher Arten. Die folgende Abbildung zeigt wie dies geschiet:
Sympatrische Artenbildung durch Hybridiserung zweier unterschiedlicher Arten (Quelle: http://wps.pearsoncustom.com)
1)
Haploide Keimzellen von zwei verschiedenen Arten verschmelzen. Der
entstandene Hybrid ist normalerweise steril, da sich seine chromsomen
aufgrund mangelnder Homologie während der Meiose nicht paaren können.
2) Jedoch kann der entstandene Hybrid sich vielleicht asexuell fortpflanzen (wie sehr viele Pflanzen können).
3)
Fehler bei der Zellteilung führen gegebenenfalls zu einer
Chromosomenverdopplung, welche zu einer polyploiden, fruchtbaren Art
führt. Auch diese Art (4n) ist von den Elternarten (2n) reproduktiv
isoliert.
Auch bei Tieren kommt es gelegentlich zu Polyploidiserung, jedoch deutlich seltener als bei Pflanzen. Ein Beispiel ist der graue Baumfrosch (siehe Bild), dessen Art sich so gebildet hat.
Meistens entsteht bei Tieren sympatrische Artenbildung jedoch eher durch die anderen, vorgestellten reproduktiven Isolationsformen
Durch Polyploidisierung entstanden: der graue Baumfrosch (Quelle: http://wps.pearsoncustom.com)
Allopatrische
Artbildung
Allopatrische Artbildung , also die Artenentstehung aufgrund geographischer Isolation gilt als einer der Hauptgründe für das Entstehen neuer Arten und setzt eine geografische Separation
von mindestens zwei Teilpopulationen voraus. Gründe für diese Art der geografischen Separation können zum Beispiel sein:
Kontinentaldrift
Gebirgsbildung
Klimawandel (z.B. Wüstenbildung o. Veränderung des Meeresspiegels)
Zufälliges abdriften in Isolate (z.B. Inseln, siehe Darwinfinken)
Sinkt beispielsweise der Wasserspiegel in einem See, kann hierdurch eine Landbrücke entstehen, die den großen See in mehrere kleinere Seen unterteilt, wodurch Fischpopulationen gespalten werden.
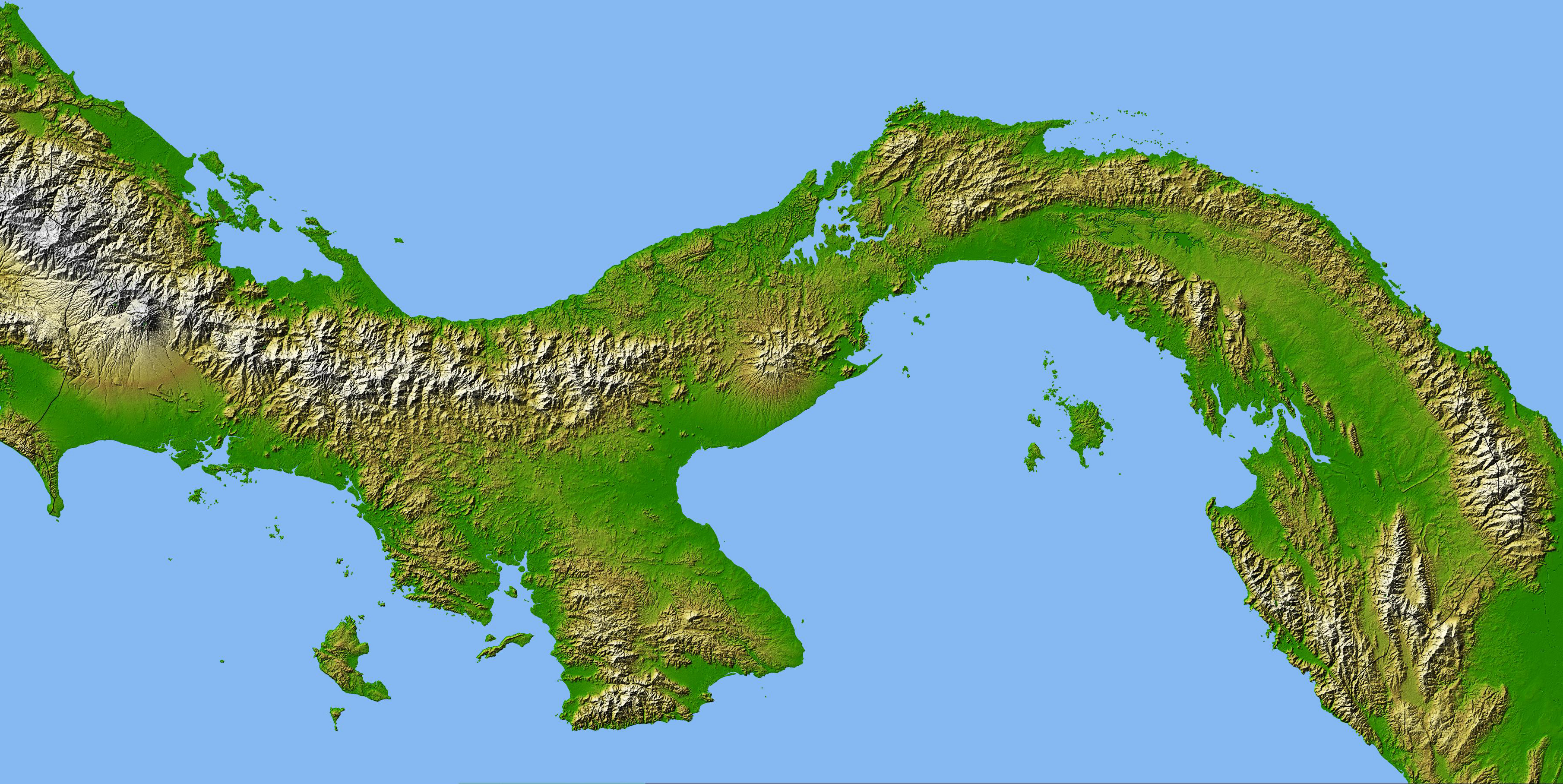
Ähnlich bildete sich vor etwa 4,5 Millionen beispielsweise auch durch Kollision zweier Kontinentalplatten der Isthmus von Panama, eine Landenge, durch die marinen Populationen auf beiden Seiten getrennt wurden. Aber auch auf andere, weniger spektakuläre Weisen können Populationen in
zwei Teilpopulationen getrennt werden. Beispielsweise kann durch einen
starken Sturm ein Teil eines Vogelschwarms abgetrennt und auf eine ferne
Insel verweht werden.
Isthmus von Panama (Quelle: earthobservatory.nasa.gov)
Da fortan kein Genfluss (keinerlei Möglichkeit Allelen auszutauschen)
mehr zwischen den beiden Populationen besteht, entwickeln
sich die Teilpopulationen aufgrund von unterschiedlichen
Selektionsfaktoren, Mutationen und auch unterschiedlichen Allelen im
Genpool auseinander. Die Selektionsfaktoren wirken anders, weil sich die
Umweltbedingungen in beiden Gebieten nicht gleichen.
Mutationen treten zufällig auf und betreffen deshalb nicht beide
Populationen gleichermaßen. Und zuletzt sorgt der Flaschenhalseffekt für
eine ungleiche
Verteilung der Allele im Genpool. Es ist denkbar, dass bestimmte
Allele nur in einer der beiden Teilpopulationen vorhanden sind.
Allopatrische Artenbildung einer Kaninchenpopulation (Quelle: u-helmich.de)
Adaptive Radiation
Unter adaptiver Radiation versteht man die Entstehung vieler neuer Arten aus einer einzigen Stammart heraus. Zur adapativen
Radiationen kommt es durch Einnischung der Art in unterschiedliche ökologische Nischen. Faktoren wie geografische Isolation,
sowie das Fehlen von natürlichen Feinden begünstigen den Vorgang der Adaptive Radiation.
Die adaptive Radiation anhand der Darwinfinken
Die Darwinfinken sind ein absolutes Musterbeispiel wenn es um die Erklärung einer adaptiven Radiation geht.
Insgesamt gibt es 14 nah verwandte Arten, die allesamt von einem gemeinsamen Vorfahren abstammen.
Auffallend sind vor allem die unterschiedlichen Schnäbel der Darwinfinken, die auf unterschiedliche Ernährungsgewohnheiten
hinweisen.
Darwin Finken(Quelle: Wikipedia)
Die Hauptnahrungsquelle des Geospiza magnirostris (1) sind Samen, während der Certhidea olivacea (4) ein
Insektenfresser ist. Dieses Prinzip der Konkurrenzvermeidung durch das Anpassen an unterschiedliche ökologische
Nischen wird gleich noch genauer erläutert.
Die Galapagosinseln liegen etwa 1000 km westlich von Südamerika entfernt und sind demnach geografisch vom Festland isoliert.
Galapagos Inseln(Quelle: Wikipedia)
Als Insel vulkanischen Ursprungs können sich die Darwinfinken auch nicht auf der Insel entwickelt haben, sondern müssen
ihren Ursprung vom Festland haben. Durch Zufall, beispielsweise durch einen Sturm oder durch Treibholz, müssen
mindestens zwei Finken (männlich und weiblich) oder ein befruchtetes Weibchen auf die Insel gelangt sein und
bildete(n) so eine Gründerpopulation. Zunächst vermehrte sich die Singvogelart sehr stark, weil neben dem
übermäßigen Nahrungsangebot auch keinerlei Fressfeinde auf der Insel vorhanden waren. Irgendwann erhöht
sich jedoch der Druck der intraspezifischen Konkurrenz auf die Finken, weil Lebensraum und Nahrung nur
begrenzt zur Verfügung stehen. Geografische Separation sorgt nun für die Entwicklung der Stammart in
unterschiedliche Richtungen. Von der Ursprungsinsel gelangen einzelne
Individuen erneut
durch Zufall auf eine weitere Insel. Der Prozess der rasanten Vermehrung wiederholt sich,
nun jedoch unter anderen Voraussetzungen, denn nicht nur der Selektionsdruck ist auf dieser anderen Insel anders (z.B. andere
Nahrungsangebote), sondern auch der Genpool unterscheidet sich nun, wenn auch nur im geringen Maße, von dem der
Ursprungspopulation der Ursprungsinsel (Flaschenhalseffekt). Des Weiteren führen unterschiedliche Mutationen und
Rekombinationen, eine veränderte Richtung der Selektion, sowie Gendrift zu einer nicht konvergenten Entwicklung wie
vergleichsweise die der Stammart auf der Ursprungsinsel.
Da zwischen den Inseln kein Genfluss mehr besteht, können die Singvögel
auch keinerlei Allele mehr miteinander austauschen
und es kommt zur Separation der beiden Populationen. Sind die beiden
Populationen lange genug getrennt, kann es in diesem
Prozess der Separation zu einer reproduktiven Isolation kommen, sodass
sich die beiden Populationen nicht mehr untereinander
fortpflanzen können. Gelangen nun Individuen der neu entstandenen Art
zurück auf die Ursprungsinsel, kann dies dazu führen,
dass diese an die selbe ökologische Nische angepasst sind und demnach
mit der anderen Art um diese ökologische Nische
konkurrieren. Nach dem Konkurrenzausschlussprinzip kann nur eine Art
eine ökologische Nische besetzen und es kommt entweder
zum Aussterben einer Art, oder eine Art ist im Stande auf eine andere
ökologische Nische auszuweichen und neben der anderen
Art zu koexistieren. Es kann jedoch auch sein, dass die neu entstandene
Art sich auf der anderen Insel derart entwickelt hat,
dass sie unmittelbar eine freie ökologische Nische auf der
Ursprungsinsel besetzt und nicht mit der Stammart in Konkurrenz tritt.
Isolierte Inselketten mit unterschiedlichen Lebensräumen (Habitaten) sind oft Schauplätze solcher adaptiver Radiationen. Die eingewandernten Kolonien durchlaufen zahlreiche allopatrische und sympatrische Prozesse, die zu Arten führen, die nirgends sonst auf der Welt gefunden werden können.
Neben den Darwinfinken , die das typische Beispiel einer adaptiven Radiation darstellen, gibt es fossile Hinweise, dass nach dem Dinosauriersterben vor 65 Mio Jahren die Säugetiere eine dramatische, adaptive Radiation erfahren haben.
Unter dem Vorgang der Mitose versteht man die einfache Zellkernteilung, bei der am Schluss
aus einer Zelle, zwei identische Tochterzellen entstehen. Funktion der Mitose ist dementsprechend die Vermehrung von Zellen.
Bei Mensch und Tier werden praktisch laufend neue Zellen gebildet. Ob bei Wachstum, Regeneration von
Verletzungen oder dem Austausch alter Zellen ist die Mitose für die Bildung neuer Zellen verantwortlich.
Bevor
es zur Mitose kommt, wird die DNA in der S-Phase repliziert, d.h. sie
wird verdoppelt, wonach jedes Chromosom aus zwei Schwester-Chromatiden
besteht.
Die Mitose ist streng von der Meiose zu unterscheiden, bei der es zu einer Reduktion der Chromosomenanzahl kommt.
(Quelle: www.biologie-schule.de)
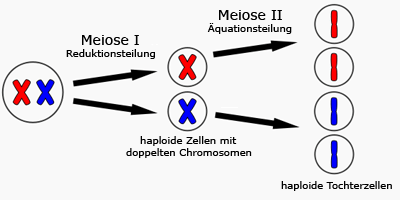
2) Meiose
Schaut man sich das Karyogramm einer Ei- oder Samenzelle an, erkennt man das diese lediglich
aus 23 Chromosomen bestehen. Alle anderen Zellen des Menschen wie z.B. Hautzellen bestehen
dagegen aus 46 Chromosomen. Dies hat einen besonderen Grund: Bei Verschmelzung von Ei- und
Sammenzelle kommt es auch zu einer Verschmelzung des Chromosomensatzes.
Die befruchtete Eizelle (sog. Zygote) besitzt dann wieder einen diploiden (doppelten) Chromosomensatz.
Keimzellen (Gameten), darunter fallen Spermien und Eizellen, besitzen nur einen haploiden
(einfachen) Chromosomensatz.
Die Meiose (griech. meiono = vermindern) ist für die Halbierung des
diploiden, auf einen haploiden Chromosomensatz bei Keimzellen zuständig.
Würde es nicht zu dieser Reduktion kommen, würde sich die Anzahl der
Chromosomen bei der Befruchtung
immer wieder addieren und es käme zu einer schier unendlichen Zahl an
Chromosomensätzen. Diese dabei entstehenden
Zygoten wären nicht lebensfähig.(Quelle: www.biologie-schule.de)
Meiose (Quelle: www.biologie.schule.de)
Evolutionsbestimmende Faktoren
Als Evolutionsfaktor bezeichnet man in der Biologie Prozesse, durch die der Genpool – das ist die Gesamtheit aller Genvariationen in einer Population – verändert wird. Eine Definition des Begriffs ergibt sich durch die Populationsgenetik: Evolutionsfaktoren sind Prozesse, die zu Veränderungen der Allelfrequenzen im Genpool einer Population führen.
Die wesentlichen Evolutionsfaktoren sind
Mutation,
Rekombination,
Selektion und
Gendrift.
1) Mutationen
Unter einer Mutation (lat. mutare = ändern) versteht man die Veränderung des Erbguts. Zunächst einmal müssen drei verschiedene Arten der Mutationen unterschieden werden.
Genmutation: Veränderung eines einzelnen Gens
Chromosomenmutation: Veränderung der Chromosomenstruktur
Genommutation: Veränderung der Anzahl der Chromosomen
Mutationen haben zwei Merkmale. Sie treten zufällig und ungerichtet auf. Das bedeutet, dass ihr Auftreten
keinen direkten Zweck verfolgt.
Außerdem treten Mutationen zeitlich spontan auf. Die Wahrscheinlichkeit zur Mutation kann durch sogenannte
Mutagene jedoch deutlich erhöht werden. Darunter zählen chemische Stoffe, Radioaktivität, UV-Strahlung und
auch Röntgenstrahlung.
Mutation ist einer der wichtigsten Evolutionsfaktoren, denn dadurch gelangen neue
Allele in den Genpool von Population. Eine Mutation kann für ein Individuum von
Vorteil, von Nachteil oder aber unbedeutend sein.
Ist eine Mutation für ein Individuum von Vorteil (z.B. verbesserte Wasseraufnahme
eines Kaktus in der Sahara oder Laktosetoleranz beim Menschen), hat es den anderen
Individuun gegenüber, die diese Mutation nicht besitzen, einen Vorteil. Im
Regelfall wird sich diese Mutation dann innerhalb der Population verbreiten, weil
eine erhöhte Fitness dazu führt, dass die Gene des Individuums häufiger in die
nächste Generation gebracht werden. Auf diese Weise breiten sich Mutationen in der
Population aus.
Ist eine Mutation jedoch von Nachteil, kann dies im schlimmsten Fall den Tod
bedeuten, etwa wenn lebenswichtige Stoffwechselvorgänge beeinflusst werden.
Mutationen die weder positive noch negative Folgen haben werden auch stille
Mutationen genannt. Denn eine Mutation kann auch an Orten in der DNA passieren, die
keinerlei wichtige Informationen enthalten oder aber die Mutation verändert nicht
die Aminosäurenabfolge im genetischen Code, weil viele Aminosäuren unterschiedliche
Codierungen zulassen.
Davon zu unterscheiden sind die neutralen Mutationen. Dort kann es zwar zu einer
phänotypischen Ausprägung kommen, diese ist dann jedoch nicht von Bedeutung für die
Selektion. Blaue Augen beim Menschen etwa sind ein Beispiel dafür. Diese Mutation
ist etwa vor 10.000 Jahren aufgetaucht.
(Quelle: www.biologie-schule.de)
2) Rekombination
Unter Rekombination versteht man die Neuverteilung von Erbgut während der Meiose.
Die Rekombination macht es quasi unmöglich, das zwei identische Nachkommen gezeugt werden und
ist somit maßgeblich für eine hohe genetische Variabilität.
Im Gegensatz zum Evolutionsfaktor Mutation, die neue Variationen schafft, sorgt die
Rekombination nur für eine Andersverteilung des vorhandenden (!) genetischen Materials.
Damit findet keine Veränderung des Genpools statt.
Hierbei sind zwei unterschiedliche Rekombinationsereignisse zu unterscheiden.
1) Intrachromosomale Rekombination
Vor der Meiose kommt es zunächst zu einer normalen Verdopplung der DNA, so dass alle Chromosomen mit zwei Chromatiden vorliegen. Während der Meiose lagern sich in der Prophase I die zwei homologen Chromosomen, also die sich jeweils entsprechenden mütterlichen und väterlichen, aneinander an: Zwischen ihnen bildet sich der Synaptonemale Komplex aus. Zwischen den mütterlichen udn väterlichen Chromatiden kann es anschliessend zu einem Austausch kommen. Dabei legen sich die Chromatiden
übereinander (crossing over = übereinander legen). Es kann dabei zu einem Bruch von Teilabschnitten kommen, die
anschließend mit Teilen des anderen Chromatids wieder geschlossen werden. Somit kommt es zu einem partiellen Austausch der Chromosomen von väterlichen und
mütterlichen Chromosomen.
Intrachromosomale Rekombination durch Crossing Over (Quelle: biologia2oppimispvkirja.blogspot.com)
2) Interchromosomale Rekombination
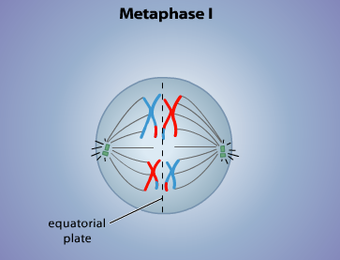
In der Metaphase innerhalb der Meiose "versammeln"
sich alle Chromosomen in der Äquatorialebene. In der folgenden
Anaphase kommt es nun zu einer zufälligen Verteilung der homologen (und zuvor rekombinierten) Chromosomen, die
von den Zugfasern des Spindelapparates an den Rand der Zelle gezogen werden. Auf diese
Weise werden die Chromosomenpaare neu kombiniert. So können sie nun aus väterlichen und
mütterlichen Chromosomen bestehen.
Metaphase I : Neue Kombination von rekombinierten, väterlichen und mütterlichen Chromosomen (Quelle:http://allaboutmeiosis.weebly.com)
3. Selektion
Die Selektion
ist die natürliche Auslese durch die Umwelt. Eine Voraussetzung für
Selektion ist die durch Rekombination und Mutation verursachte
Variabilität in einer Population. In den meisten Populationen werden
viel mehr Nachkommen erzeugt, als in ihrem Lebensraum überleben können.
So sterben viele Individuen einer Generation, bevor sie sich
fortpflanzen können, oder bekommen weniger Nachkommen als andere
Individuen mit vorteilhafteren Merkmalsausprägungen. Die Individuen
leisten also einen unterschiedlichen Beitrag zum Genpool der
Folgegeneration. Die relativen Häufigkeiten der Erbanlagen mit günstigen
Merkmalsausprägungen nehmen in der Population zu, während die relativen
Häufigkeiten der Erbanlagen mit ungünstigen Merkmalsausprägungen
abnehmen.
Man unterscheidet drei Formen von Selektion:
Natürliche Selektion
An ihre Umwelt besser angepasste Lebewesen haben eine höhere
Wahrscheinlichkeit ihre Gene zu vererben als Lebewesen mit einer schlechter angepassten Lebewese.
Hierunter versteht man die innerartliche Auswahl von Sexualpartnern,
die sich aus der Konkurrenz um Fortpflanzungspartner ergibt. Sexuelle Selektion erklärt auch zahlreiche phänotypische Ausprägungen, die im Sinne der natürlichen Selektion eigentlich von Nachteil
wären (z.B. das Federkleid des Pfau. Dieses ist hinderlich bei der Flucht vor Räubern, ist
aber wichtiges "Balzmittel").
Pfau (Quelle: www.natur-lexikon.de)
Künstliche Selektion
Unter künstlicher Selektion versteht man die vom Menschen gesteuerte
Selektion zur Förderung bestimmter (erwünschter) Merkmale bei Tier- und Pflanzenarten. Hierunter fallen zum Beispiel eine höhere
Milchleistung von Kühen oder eine höhrere Resistenz von Nutzpflanzen gegenüber Krankheiten und Umweltbedingungen. Unter künstliche Selektion fallen die Pflanzenzucht sowie die Kleintierzucht.
Unter Selektionsfaktoren versteht man jene Umweltbedingungen/faktoren, die auf die
Individuen, und damit auf deren Fitness einwirken. Dabei wird zwischen abiotischen
(unbelebten) und biotischen (belebten) Selektionsfaktoren unterschieden.
Abiotische Selektionsfaktoren
Unter den abiotischen Selektionsfaktoren fasst man die Faktoren zusammen, die von einer unbelebten Umwelt ausgehen. Hierunter fallen Parameter wie Temperatur, Feuchtigkeit, Wind, Nährstoffe, oder Gifte.
Beispiel Temperatur
Im Hinblick auf die Temperatur als abiotischer Selektionsfaktor spielen die sogenannten Klimaregeln eine wichtige Rolle:
1. Bergmannsche Regel
Die Bergmannsche Regel ist die sog. erste Klimaregel, benannt nach C. Bergmann.
Die meist nur auf Warmblüter anwendbare Bergmannsche Regel beschreibt
die Beobachtung, daß innerhalb einer Art die Individuen von Populationen
aus kalten Gebieten größer sind als in den warmen. Bergmann erklärte
dies mit den unterschiedlichen Verhältnissen zwischen der Oberfläche und
dem Volumen bei kleinen bzw. großen Körpern. Ein großer Körper verliert
über seine (in bezug auf sein Volumen) relativ geringe Oberfläche
weniger Wärme als ein kleiner. Entsprechend sind nördlich verbreitete
Populationen mancher Säugetiere (z. B. Hirsch, Wildschwein) und Vögel (z. B. Uhu, Gimpel) größer als südliche (Quelle: www.spektrum.de) .
Bergmann´sche Regel (Quelle: www.biologieunterricht.de)
2. Allensche Proportionsregel
Die Allensche Proportionsregel ist benannt nach dem amerikan. Zoologen Joel Asaph Allen, 1838–1921. Sie ist die
2. Klimaregel, die besagt, daß bei verwandten Säugetier-Arten oder
-Rassen die exponierten Körperteile (wie Schwänze und Ohren) in kalten
Gebieten relativ kürzer ausgebildet sind als in wärmeren Gebieten.
Beispiele: der Eisfuchs mit sehr kleinen Ohren, der Rotfuchs (Füchse) mit mittelgroßen und der Wüstenfuchs oder Fennek mit sehr großen Ohren. Die mächtigen Ohren des Afrikanischen Elefanten (Gesamtoberfläche 8 m2)
dienen geradezu der Wärmeregulation und werden bei großer Hitze weit
abgestellt und durch Bewegung ventiliert (bei niedrigeren Temperaturen
und bei Regen hingegen ruhig gehalten und dem Körper angelegt); die des
Indischen Elefanten sind auffallend kleiner; besonders kleine Ohren aber
hatte das Mammut, das in der Eiszeit die kalten Steppen und Tundren der Nordhalbkugel bewohnte. (Quelle: www.spektrum.de)
Allen´sche Regel(Quelle: www.scienceblogs.de)
Allen´sche Regel am Beispiel verschiedener Fuchsarten (Quelle: www.biologieunterricht.de)
Bergmann´sche und Allen´sche Regel (Quelle: www.weserkurier.de)
Beispiel Feuchtigkeit
Außergewöhnliche Umweltbedinungen, wie etwa die enorme
und langanhaltende Trockenheit in Wüstengebieten, führen zu Pflanzen mit sehr
geringem Wasserverbrauch, bis hin zur Ausbildung von sogenannten Sukkulenten, also besonders saftreiche Pflanzen (Quelle: www.biologie-unterricht.de).
Die Flügel der Kerguelenfliege (bewohnen die Kerguelen
Inselgruppen im Indischen Ozean) haben sich im Laufe der Evolution zu Stummeln
zurückgebildet. Fliegen mit ausgebildetem Flügelpaar wurden häufig durch Stürme auf
das offene Meer herausgeweht. Auf windigen Inseln kann es daher ein evolutionärer
Vorteil sein, verkümmerte Flügel zu besitzen (Quelle: www.biologie-unterricht.de).
Kerguelen(Quelle: http://unimog-club-gaggenau.de)
Die Kerguelen Fliege (Quelle: http://www.cosmiq.de/)
Beispiel Nährstoffe
Das Nährstoffangebot stellt einen typischen abiotischen Selektionsfaktor dar.
Fleischfressende Pflanzen sind beispielsweise in der Lage auf
stickstoffarmen Böden zu siedeln, da sie die fehlenden Mineralstoffe über ihre
Beutetiere ausgleichen können (Quelle: www.biologie-unterricht.de).
Venusfliegenfalle (Quelle: www.allmystery.de)
Beispiel Gifte
Der Einsatz von Antibiotika wirkt als Selektionsdruck, der die Evolution resistenter Bakterien fördert.
Unter biotischen Selektionsfaktoren werden sämtliche Selektionsfaktoren zusammengefasst, die von der belebten Umwelt
ausgehen. Dabei können zwischen Intraspezifischen (innerartlich) und
Interspezifischen (außerartlich) Faktoren unterschieden werden (Quelle: www.biologie-unterricht.de).
Intraspezifische Selektionsfaktoren
Hierunter fallen die bereits besprochenen Formen der sexuellen Selektion, also die Merkmalsausprägungen, die zu einer höheren, innerartlichen Fortpflanzungs-wahrscheinlichkeit führen. Beispiele sind das Federkleid mancher Vogelarten (Pfau) oder das Hirschgeweih.
Das Hirschgeweih als ein intraspezifischer Selektionsfaktor (Quelle: www.tiergaerten.de)
Interspezifische Selektionsfaktoren
Interspezifische Selektionsfaktoren sind in bestehenden Räube-Beute Beziehungen begründet und äussern sich in Imitierungen anderer Lebewesen oder der Umwelt. Hierdurch sind die entsprechenden Lebewesen dann entweder besser an die Umgebung angepasst und entweder für Beutetiere und/oder Freissfeinde nicht so leicht zu entdecken (z.B. wandelndes Blatt, Lebende Steine, Chamäleon), oder aber gefährliche Tiere werden imitiert, um so den Eindruck zu erwecken, selbst gefährlich zu sein (Mimikry). Beispiel: Die harmlose Schwebfliege sieht der Wespe zum verwechseln ähnlich, und
wird so von potentiellen Fressfeinden gemieden.
Wandelnde Blatt (Quelle: www.geo.de)
Lebende Steine (www.gartenforum.de)
Schwebfliege (Quelle: www.blackstein,de)
4) Gendrift
Unter Gendrift versteht man die zufällige Veränderung der Genhäufigkeit eines bestimmten Allels innerhalb einer
Population. Besonders bei kleinen Populationen ist der Gendrift bedeutend, weil Allele relativ schnell aus dem
Genpool der Population verschwinden können. Aber auch das genaue Gegenteil ist möglich, nämlich dass bestimmte
Gene plötzlich extrem häufig in einer Population auftauchen, etwa nach Naturkatastrophen, wenn nur noch wenige
Individuen überlebt haben und viele von ihnen ein zuvor noch seltenes Allel in sich tragen (Quelle: www.biologie-schule.de). Es gibt zwei bekannte Formen von Gendrift:
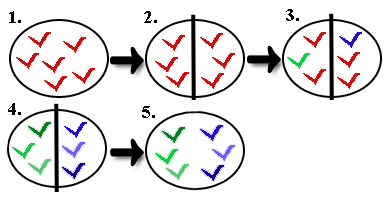
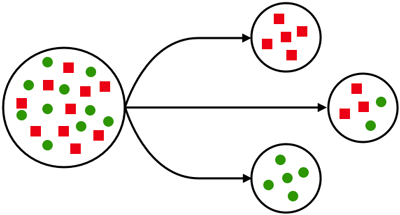
1) Gründer-Effekt
Beim Gründereffekt wird eine neue Population durch einige wenige Individuen der Ausgangspopulation (P1) gegründet. Innerhalb dieser neuen
Gründerpopulation (P2) unterscheidet sich die Allelfrequenz (Allelhäufigkeit) im Vergleich zur nun
isolierten Ausgangspopulation (P1). Es besteht kein Genfluss mehr zwischen P1 und P2. Dies hat eine
deutlich geringere Genvariabilität zur Folge, weil manche Allele, die in P1 auftauchen, in keinem der
Individuen von P2 vorhanden sind. Der Gründereffekt kann somit zum leichteren Aussterben
kleiner, isolierter Populationen beitragen. Viele Tier- und Pflanzenarten, die in
Restpopulationen im Freiland überleben oder in Erhaltungszuchten
gerettet werden, unterliegen dem Gründereffekt.
Notwendige Bedingung des Gründereffekts ist Isolation von der Ausgangspopulation, etwa durch geografische
Isolation.
Gründereffekt ( Quelle: www.scilogs.de)
Gründereffekt
2) Flaschenhalseffekt
Der Flaschenhalseffekt beschreibt die starke Reduzierung der genetischen Variabilität in Verbindung mit der
randomisierten Änderung der Allelhäufigkeiten. Dem Flaschenhalseffekt liegt meist ein Ereignis zu Grunde, etwa eine
Naturkatastrophe in der viele Individuen getötet werden oder plötzliche geografische Isolation durch Plattentektonik. In allen Fällen ist die zufällige Auswahl der
Individuen für den Flaschenhalseffekt entscheidend, unabhängig von ihrer Anpassung an die Umweltfaktoren. Beim Erhalt stark gefährdeter Arten kann dies ein zentrales Problem darstellen, wenn ein die Reproduktion beeinträchtigendes Allel nicht durch ein zweites, für die Reproduktion vorteilhafteres Allel ausgeglichen werden kann.
Flaschenhalseffekt
Der Unterschied zwischen Flaschenhals- und Gründereffekt
Der Unterschied zwischen dem Flaschenhals- und Gründereffekt ist die
Art des Zustandekommens. Beim Flaschenhalseffekt hast du ursprünglich
eine Population (breite Basis); dann kommt es zu einer Umweltveränderung
(zum Beispiel eine Katastrophe oder die Veränderung eines Flusslaufs),
was entweder den Großteil der ursprünglichen Poulation vernichtet oder
einen kleinen Teil vom Rest abschneidet (schmaler Flaschenhals). Im
überlebenden (separierten) Teil kann der Genpool nun von dem der
ursprünglichen Population mehr oder weniger stark abweichen. Dadurch
haben plötzlich auch Genotypen eine Chance zur Fortpflanzung, die ohne
die Veränderung sich nicht durchgesetzt hätten.
Beim Gründereffekt werden einzelne Individuen in separierte Refugien
verschlagen, zum Beispiel treiben auf einem Stück Holz sehr wenige
Individuen vom Festland zu einer Insel. Auf der Insel angekommen,
vermehren sich die Tiere und bilden so den Ausgangspunkt für eine neue
Population (Gründerindividuen). Es liegt auf der Hand, dass der Genpool
der neu entstehenden Inselpopulation ausschließlich von der genetischen
Ausstattung der Gründer abhängt.
Von der Evolution aus betrachtet, sind beide Effekte einander sehr
ähnlich, weil es in beiden Fällen zu erheblichen Allelverschiebungen
kommen kann (verglichen mit dem ursprünglichen Genpool), so dass sich
neue Arten mit anderen Merkmalen herausbilden können, die ohne die
Effekte nicht entstanden wären (weil sie möglicherweise objektiv
Selektionsnachteile haben, die sich nur ausprägen können, weil es keine
Alternative gibt...) (Quelle: www.gutefrage.net).