Mechansimen der Artentstehung

Der Artbegriff
Als Arten bezeichnet man Gruppen von Individuen, die durch Abstammungsbande zwischen Elter(n) und Nachkommen (Nachkommenschaft) gekennzeichnet sind und in Gestalt, Physiologie und Verhalten soweit übereinstimmen, daß sie sich von anderen Individuengruppen abgrenzen lassen. Bei Organismen mit zweigeschlechtlicher Fortpflanzung kommt als entscheidendes Kriterium die Fähigkeit hinzu, gemeinsam fertile Nachkommen zu erzeugen. Die Angehörigen einer Art bilden demnach eine Fortpflanzungsgemeinschaft, zwischen ihnen besteht Genfluß, sie haben Anteil an einem Genpool und sind somit die Einheit, in der evolutionärer Wandel stattfindet (Evolution). – In aller Regel lassen sich die Individuen verschiedener Biospezies anhand äußerer Merkmale unterscheiden, doch liefern nicht diese, sondern allein die zu beobachtenden Fortpflanzungsbeziehungen das entscheidende Kriterium artlicher Zuordnung. Dies zeigt sich vor allem in kritischen Fällen, z. B. bei stark ausgeprägtem Sexualdimorphismus (wo man die beiden Geschlechter primär unterschiedlichen Arten zuordnen würde) oder bei den morphologisch sehr ähnlichen, aber genetisch isolierten Zwillingsarten (die man fälschlich als eine Art auffassen kann) (Quelle: www.spektrum.de).Reproduktive Isolation
Die reproduktive Isolation bildet die essentielle Grundlage der Artentstehung. Bei einer reproduktiven Isolation besteht durch eine oder mehrere Fortpflanzungsbarrieren kein Genfluss mehr.
Fortpflanzungsbarrieren können innerartlich, aber auch außerartlich wirken und führen dauerhaft zum Entstehen einer neuen
Art (Ausnahme: Isolation durch Sterilität).
Man unterscheidet mehrere Formen von Isolation:
Zeitliche Isolation
Arten können sich nicht miteinander fortpflanzen, weil sie sich während unterschiedlicher Jahreszeiten/Tageszeiten fortpflanzen
(z.B. bei Froscharten, deren Paarungszeit in unterschiedlichen Monaten liegt)
Habitat Isolation
Zwei eng verwandte Arten von Strumpfbandnattern paaren sich nicht, da eine Art an Land lebt während die Andere hauptsächlich im Wasser. |
| Habitat Isolation zweier Stumpfbandnatter-Arten (Quelle: http://wps.pearsoncustom.com) |
Genetische Isolation
Durch zufällige Mutationen können Individuen nicht mehr mit der
Ursprungspopulation fortpflanzungsfähig sein.
Physiologische Isolation
Aufgrund der unterschiedlichen Form der Kopulationsorgane können sich bestimmte Arten nicht miteinander fortpflanzen
(z.B. bei nah verwandten Insektenarten).
Bei den hier gezeigten verschiedenen Schneckenarten drehen die Schneckenhäuser in entgegengesetzten Richtungen. Hierdurch können die Genitalöffnungen )siehe Pfeile) nicht in die richtige Position gebracht werden und somit keine Paarung durchgeführt werden.
Bei den hier gezeigten verschiedenen Schneckenarten drehen die Schneckenhäuser in entgegengesetzten Richtungen. Hierdurch können die Genitalöffnungen )siehe Pfeile) nicht in die richtige Position gebracht werden und somit keine Paarung durchgeführt werden.
 |
| Physiologische Isolation bei Schnecken (Quelle: http://wps.pearsoncustom.com) |
Ökologische Isolation
Durch das Ausnutzen von unterschiedlichen ökologischen
Nischen im selben Gebiet kommt es zu einer reproduktiven Isolation (z.B. Darwinfinken, die unterschiedliche ökologische Nischen
besetzten; manche Individuen ernährten sich von Insekten, andere von Körnern).
 |
| Ökologische Isolation bei Darwinfinken |
Geografische Isolation
Wegen geografischen Barrieren können Teilpopulationen einer Art sich nicht untereinander fortpflanzen
(z.B. durch Kontinentaldrift oder eine Erhöhung des Meeresspiegels).
Verhaltensisolation
Unterschiedliches Verhalten während der Paarungszeit isoliert Arten voneinander (z.B. haben nah verwandte Vogelarten, die sich zur selben Zeit paaren, einen unterschiedlichen Balzruf).Sterilität
Bei der Kreuzung zweier nicht verwandten Arten können Bastarde mit einem ungeraden Chromosomensatz entstehen, die nicht Fortpflanzungsfähig sind (z.B. bei einer Kreuzung aus Esel und Pferd sind die Nachkommen allesamt unfruchtbar, weil sie einen ungeraden Chromosomensatz besitzen, der eine Bildung von Gameten unmöglich macht). |
| Die Kreuzung von Pferd und Esel bringt Nachkommen mit einem ungeraden Chromosomensatz hervor, die nicht fortpflanzungsfähig sind. (Quelle: http://wps.pearsoncustom.com) |
Polyploidie
Unter Polyploidie versteht man das Vorhandensein von mehr als zwei Chromosomensätzen. Fortpflanzung ist nur unter Individuen mit identischer Anzahl von Chromosomensätzen möglich. (z.B. bilden Pflanzen häufig triploide oder tetraploide Chromosomensätze und sind somit von ihrer Ursprungsart mit haploidem Chromosomensatz genetisch isoliert).
(Quelle: www.biologie-heute.de,
http://wps.pearsoncustom.com)
Fortpflanzungsbarrieren als Resultat der Isolierung
Bei einer Isolation können sich Fortpflanzungsbarrieren entwickeln. Die zeigt das folgende Experiment von Diane Dodd: Eine Population von Fruchtfliegen wurde aufgeteilt und fortan für mehrere Generationen entweder in Anwesenheit von Maltose oder Stärke als Kohlenstoffquelle kultiviert. Anschliessend wurde ein Paarungsexperiment durchgeführt, und die Paarungsversuche statistisch ausgewertet. Das Experiment zeigt, dass sich die Fliegen, die Stärke als Kohlenstoffquelle gewohnt sind, auch präferentiell mit solchen Fliegen paaren. Ebenso verhalten sich die Fliegen, die Maltose als Kohlenstoffquelle gewohnt sind. Zwar treten auch Paarungen zwischen den Gruppen auf (Stärke-Maltose) , jedoch sind diese deutlich seltener (< 50%) .
 |
| Diane Dodd: Als Folge von isolation entstehen Fortpflanzungsbarrieren (Quelle: http://wps.pearsoncustom.com) |
 |
| Isolation von Populationen kann zu Fortpflanzungsbarrieren führen (Quelle: UC Berkeley) |
Sympatrische Artenbildung
Die Entstehung einer neuen Art im Gebiet der Ursprungsart nennt man sympatrische Artbildung. Bei dieser Form der Artbildung
muss es, beispielsweise durch Polyploidisierung bei Pflanzenarten, zu
einer unmittelbaren reproduktiven Isolationen kommen, damit
die Pflanze keinen Allelaustausch mehr mit der übrigen Population hat.
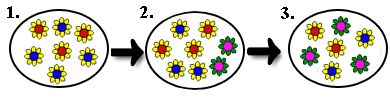 |
| Sympatrische Artenbildung (Quelle: www.biologie-schule.de) |
1) Sympatrische Artenbildung durch Polyploidisierung
Viele Pflanzenarten sind das Ergebnis eines Fehlers bei der Zellteilung, durch welchen ein zusätzlicher Satz an Chromosomen entstanden ist. Arten, die sich auf diese Weise gebildet haben sind polyploid, d.h. ihre Zellen haben mehr als zwei Chromosomensätze. In der folgenden Abbildung ist gezeigt, wie sich aus einer diploiden Pflanze (2n) eine tetraploide Pflanze entwickeln kann:
 |
| Sympatrische Artenbildung durch Polyploidisierung (Quelle: http://wps.pearsoncustom.com) |
1)
Ein Fehler bei der Zellteilung führt nach der Replikation des
diploiden Chromosomensatzes (2n) zu einer Verdopplung der Chromosomen.
2)
Wenn sich aus diesen tetraploiden Zellen (4n) eigenständige
Pflanzen entwickeln, bilden diese Pflanzen Blüten mit diploiden Gameten
(Keimzellen) aus.
3)
Wenn diese Pflanzen sich selbst befruchten (was häufig bei Pflanzen
vorkommt), entwickeln sich aus den tetraploiden Zygoten erneut
tetraploide Pflanzen, die fruchtbar sind und durch Selbstbefruchtung
oder Befruchtung durch andere tetraploide Pflanzen neue, tetraploide
Nachkommen bilden. Die so entstandenen tetraploiden Pflanzen können jedoch keine fruchtbaren Nachkommen erzeugen, wenn sie durch die Elternpflanzen befruchtet werden (und auch umgekehrt nicht !). Die Verschmelzung der diploiden Keimzellen der tetraploiden Pflanze mit den haploiden Keimzellen der diploiden Elternpflanze würde eine triploide Nachkommenschaft (3n) erzeugen. Triploide Individuen sind jedoch unfruchtbar. Sie können keine normalen Keimzellen bilden, da durch die ungerade Chromosomenzahl während der Meiose keine homologen Chromosomenpaare gebildet werden können, die sich anschliessend durch Zellteilung gleich aufteilen würden. Somit wird durch die Bildung eines tetraploiden (4n) Pflanzennachkommens innerhalb nur einer Generation eine neue Art geschaffen, die reproduktiv isoliert ist.
2) Sympatrische Artenbildung durch Hybridisierung
Die meisten polyploiden Pflanzen entwicklen sich jedoch durch Hybridisierung zweier unterschiedlicher Arten. Die folgende Abbildung zeigt wie dies geschiet:
 |
| Sympatrische Artenbildung durch Hybridiserung zweier unterschiedlicher Arten (Quelle: http://wps.pearsoncustom.com) |
1)
Haploide Keimzellen von zwei verschiedenen Arten verschmelzen. Der
entstandene Hybrid ist normalerweise steril, da sich seine chromsomen
aufgrund mangelnder Homologie während der Meiose nicht paaren können.
2) Jedoch kann der entstandene Hybrid sich vielleicht asexuell fortpflanzen (wie sehr viele Pflanzen können).

3) Fehler bei der Zellteilung führen gegebenenfalls zu einer Chromosomenverdopplung, welche zu einer polyploiden, fruchtbaren Art führt. Auch diese Art (4n) ist von den Elternarten (2n) reproduktiv isoliert.
Auch bei Tieren kommt es gelegentlich zu Polyploidiserung, jedoch deutlich seltener als bei Pflanzen. Ein Beispiel ist der graue Baumfrosch (siehe Bild), dessen Art sich so gebildet hat.
Meistens entsteht bei Tieren sympatrische Artenbildung jedoch eher durch die anderen, vorgestellten reproduktiven Isolationsformen
 |
| Durch Polyploidisierung entstanden: der graue Baumfrosch (Quelle: http://wps.pearsoncustom.com) |
Allopatrische Artbildung
Allopatrische Artbildung , also die Artenentstehung aufgrund geographischer Isolation gilt als einer der Hauptgründe für das Entstehen neuer Arten und setzt eine geografische Separation
von mindestens zwei Teilpopulationen voraus. Gründe für diese Art der geografischen Separation können zum Beispiel sein:
- Kontinentaldrift
- Gebirgsbildung
- Klimawandel (z.B. Wüstenbildung o. Veränderung des Meeresspiegels)
- Zufälliges abdriften in Isolate (z.B. Inseln, siehe Darwinfinken)
Sinkt beispielsweise der Wasserspiegel in einem See, kann hierdurch eine Landbrücke entstehen, die den großen See in mehrere kleinere Seen unterteilt, wodurch Fischpopulationen gespalten werden.
Ähnlich bildete sich vor etwa 4,5 Millionen beispielsweise auch durch Kollision zweier Kontinentalplatten der Isthmus von Panama, eine Landenge, durch die marinen Populationen auf beiden Seiten getrennt wurden.
Aber auch auf andere, weniger spektakuläre Weisen können Populationen in zwei Teilpopulationen getrennt werden. Beispielsweise kann durch einen starken Sturm ein Teil eines Vogelschwarms abgetrennt und auf eine ferne Insel verweht werden.
Aber auch auf andere, weniger spektakuläre Weisen können Populationen in zwei Teilpopulationen getrennt werden. Beispielsweise kann durch einen starken Sturm ein Teil eines Vogelschwarms abgetrennt und auf eine ferne Insel verweht werden.
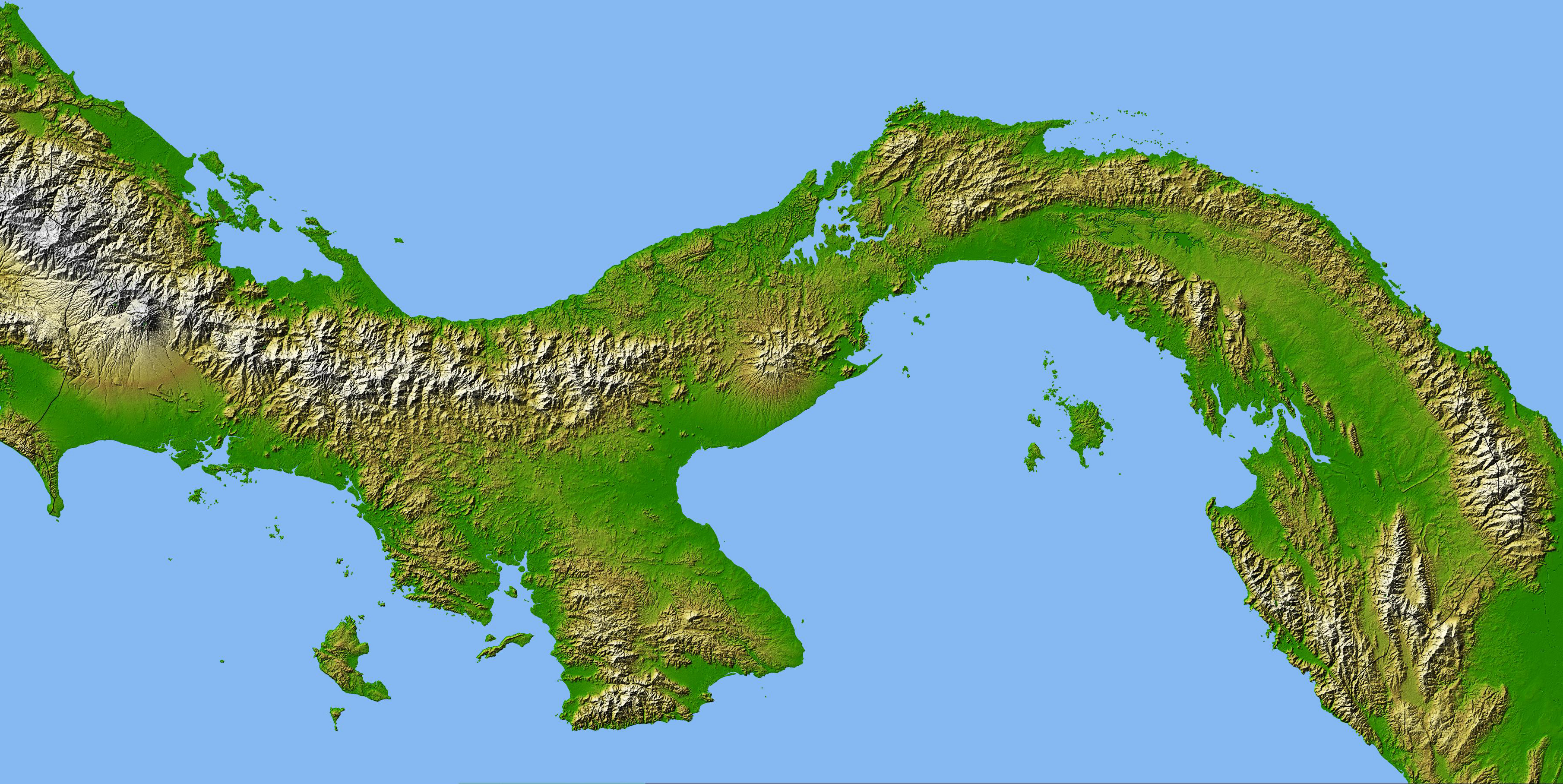 |
| Isthmus von Panama (Quelle: earthobservatory.nasa.gov) |
Da fortan kein Genfluss (keinerlei Möglichkeit Allelen auszutauschen)
mehr zwischen den beiden Populationen besteht, entwickeln
sich die Teilpopulationen aufgrund von unterschiedlichen
Selektionsfaktoren, Mutationen und auch unterschiedlichen Allelen im
Genpool auseinander. Die Selektionsfaktoren wirken anders, weil sich die
Umweltbedingungen in beiden Gebieten nicht gleichen.
Mutationen treten zufällig auf und betreffen deshalb nicht beide
Populationen gleichermaßen. Und zuletzt sorgt der Flaschenhalseffekt für
eine ungleiche
Verteilung der Allele im Genpool. Es ist denkbar, dass bestimmte
Allele nur in einer der beiden Teilpopulationen vorhanden sind.
(Quelle: www.biologie-heute.de, http://wps.pearsoncustom.com, u-helmich.de)
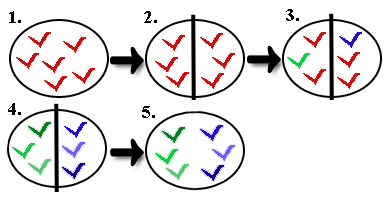 |
| Allopatrische Artenbildung (Quelle: www.biologie.schule.de) |
 |
| Allopatrische Artenbildung einer Kaninchenpopulation (Quelle: u-helmich.de) |
Adaptive Radiation
Unter adaptiver Radiation versteht man die Entstehung vieler neuer Arten aus einer einzigen Stammart heraus. Zur adapativen
Radiationen kommt es durch Einnischung der Art in unterschiedliche ökologische Nischen. Faktoren wie geografische Isolation,
sowie das Fehlen von natürlichen Feinden begünstigen den Vorgang der Adaptive Radiation.
Die adaptive Radiation anhand der Darwinfinken
Die Darwinfinken sind ein absolutes Musterbeispiel wenn es um die Erklärung einer adaptiven Radiation geht.
Insgesamt gibt es 14 nah verwandte Arten, die allesamt von einem gemeinsamen Vorfahren abstammen.
Auffallend sind vor allem die unterschiedlichen Schnäbel der Darwinfinken, die auf unterschiedliche Ernährungsgewohnheiten
hinweisen.
|
| Darwin Finken (Quelle: Wikipedia) |
Die Hauptnahrungsquelle des Geospiza magnirostris (1) sind Samen, während der Certhidea olivacea (4) ein
Insektenfresser ist. Dieses Prinzip der Konkurrenzvermeidung durch das Anpassen an unterschiedliche ökologische
Nischen wird gleich noch genauer erläutert.
Die Galapagosinseln liegen etwa 1000 km westlich von Südamerika entfernt und sind demnach geografisch vom Festland isoliert.
 | |
| Galapagos Inseln (Quelle: Wikipedia) |
Als Insel vulkanischen Ursprungs können sich die Darwinfinken auch nicht auf der Insel entwickelt haben, sondern müssen ihren Ursprung vom Festland haben. Durch Zufall, beispielsweise durch einen Sturm oder durch Treibholz, müssen mindestens zwei Finken (männlich und weiblich) oder ein befruchtetes Weibchen auf die Insel gelangt sein und bildete(n) so eine Gründerpopulation. Zunächst vermehrte sich die Singvogelart sehr stark, weil neben dem übermäßigen Nahrungsangebot auch keinerlei Fressfeinde auf der Insel vorhanden waren. Irgendwann erhöht sich jedoch der Druck der intraspezifischen Konkurrenz auf die Finken, weil Lebensraum und Nahrung nur begrenzt zur Verfügung stehen. Geografische Separation sorgt nun für die Entwicklung der Stammart in unterschiedliche Richtungen. Von der Ursprungsinsel gelangen einzelne Individuen erneut durch Zufall auf eine weitere Insel. Der Prozess der rasanten Vermehrung wiederholt sich, nun jedoch unter anderen Voraussetzungen, denn nicht nur der Selektionsdruck ist auf dieser anderen Insel anders (z.B. andere Nahrungsangebote), sondern auch der Genpool unterscheidet sich nun, wenn auch nur im geringen Maße, von dem der Ursprungspopulation der Ursprungsinsel (Flaschenhalseffekt). Des Weiteren führen unterschiedliche Mutationen und Rekombinationen, eine veränderte Richtung der Selektion, sowie Gendrift zu einer nicht konvergenten Entwicklung wie vergleichsweise die der Stammart auf der Ursprungsinsel.
Da zwischen den Inseln kein Genfluss mehr besteht, können die Singvögel auch keinerlei Allele mehr miteinander austauschen und es kommt zur Separation der beiden Populationen. Sind die beiden Populationen lange genug getrennt, kann es in diesem Prozess der Separation zu einer reproduktiven Isolation kommen, sodass sich die beiden Populationen nicht mehr untereinander fortpflanzen können. Gelangen nun Individuen der neu entstandenen Art zurück auf die Ursprungsinsel, kann dies dazu führen, dass diese an die selbe ökologische Nische angepasst sind und demnach mit der anderen Art um diese ökologische Nische konkurrieren. Nach dem Konkurrenzausschlussprinzip kann nur eine Art eine ökologische Nische besetzen und es kommt entweder zum Aussterben einer Art, oder eine Art ist im Stande auf eine andere ökologische Nische auszuweichen und neben der anderen Art zu koexistieren. Es kann jedoch auch sein, dass die neu entstandene Art sich auf der anderen Insel derart entwickelt hat, dass sie unmittelbar eine freie ökologische Nische auf der Ursprungsinsel besetzt und nicht mit der Stammart in Konkurrenz tritt.
Isolierte Inselketten mit unterschiedlichen Lebensräumen (Habitaten) sind oft Schauplätze solcher adaptiver Radiationen. Die eingewandernten Kolonien durchlaufen zahlreiche allopatrische und sympatrische Prozesse, die zu Arten führen, die nirgends sonst auf der Welt gefunden werden können.
Neben den Darwinfinken , die das typische Beispiel einer adaptiven Radiation darstellen, gibt es fossile Hinweise, dass nach dem Dinosauriersterben vor 65 Mio Jahren die Säugetiere eine dramatische, adaptive Radiation erfahren haben.